Het Anabolenboek
Willem Koert
Aede de Groot
Wageningen, 26/1/2007
8. Werking van het Complex van Testosteron en zijn Receptor
Aede de Groot, Willem Koert
De vorming van het steroid-androgeenreceptor-complex is een essentiele stap in de fysiologische processen die de androgeenreceptor regelt. Voordat het complex ontstaat, moet het lichaam het anabole steroid eerst transporteren naar de cellen waar het moet werken.
De biosynthese van testosteron vindt plaats in de testes. Het bloed zorgt daarna voor het transport naar andere plaatsen in het lichaam. Dit transport van testosteron vindt plaats in de vrije vorm en gebonden aan eiwitten [1]. Binding aan eiwitten is noodzakelijk voor een goed transport omdat de apolaire steroiden niet goed oplosbaar zijn in het polaire waterige milieu van het bloed.
Een tweede reden voor het binden van testosteron aan transporteiwitten is dat binding verhindert dat het testosteron voortijdig afbreekt. Als steroiden zijn gebonden aan eiwitten, zijn ze niet meer goed toegankelijk voor de enzymen die deze steroďden kunnen omzetten in andere, niet meer werkzame verbindingen. We zullen daar in de volgende hoofdstukken dieper op in gaan.
Slechts twee procent van de totale hoeveelheid testosteron in het bloed is aanwezig in vrije vorm. Ongeveer vijfenveertig procent is zwak gebonden aan serum albumine, een bloedeiwit. De overige vijftig tot zestig procent van het in het bloed aanwezig testosteron is stevig gebonden aan het eiwit sex hormoon bindend globuline, dat we kortweg SHBG noemen. SHBG bindt ook het vrouwelijke geslachtshormoon estradiol, zij het in mindere mate. De binding van testosteron aan serum albumine is zwak en een snelle dissociatie is mogelijk. De relatief grote hoeveelheid testosteron die in het bloed aan albumine is gebonden, kan snel overgaan naar de vrije werkzame vorm.
Alleen de vrije vorm van testosteron is biologisch actief, en kan de cel binnen dringen waar het zijn functie uitoefent. Cellen zijn omgeven door celwanden, maar dat zijn geen ondoordringbare scheidingswanden voor kleine moleculen als testosteron. Dat geldt ook voor allerlei (voedings)stoffen, die voortdurend de cel in moeten kunnen, en afvalstoffen die de cel weer moeten verlaten.
De vrije geslachtshormonen komen via diffusie uit het bloed de cel in. Dat wil zeggen dat de spontane eigen beweging van de testosteronmoleculen voldoende is om ze in de cel te brengen. Er is geen speciaal transporteiwit nodig voor dat proces.
Eenmaal in de celvloeistof aangekomen moet het testosteron molecuul zijn receptor vinden. In de cel zijn ontzettend veel stoffen aanwezig. Alles bij elkaar zijn dat ongeveer 1014-1015 moleculen. We hebben het dan over getallen met 14 tot 15 nullen. Deze moleculen zijn deels gebonden aan celmembranen, maar de meeste bewegen zich vrij in de celvloeistof. Er vinden voortdurend botsingen plaats tussen deze moleculen. Op een gegeven moment vindt de juiste botsing plaats tussen testosteron en de androgeenreceptor. Dan vindt herkenning plaats, gebaseerd op de interacties die we in hoofdstuk 6 hebben beschreven.
De kans dat testosteron botst op zijn receptor is groter wanneer er meer testosteron en meer androgeenreceptor in de cel aanwezig is.
We gaan nu eerst iets vertellen over de bouw van de androgeenreceptor. Daarna gaan we in op wat het complex van receptor en androgeen verder doet in de celvloeistof en in de celkern. We maken daarbij gebruik van goede overzichtsartikelen over die onderwerpen [2-6].
We hebben al verteld dat de androgeenreceptor een groot eiwitmolecuul is. Het is opgebouwd uit 919 aminozuren.
De aminozuurketen van het eiwit is op opgevouwen tot een grote kluwen. Aan de buitenkant en binnenin die kluwen
liggen verschillende domeinen die elk hun eigen functie hebben.
i) Het ligand-bindende domein (LBD), dat zorgt voor de vorming van de holte en de binding met de
liganden testosteron en dihydrotestosteron.
ii) Het kern-translocatie-signaal (Nuclear Location Signal of NLS), dat ervoor zorgt dat het
ligand-receptorcomplex het membraan dat de celkern omgeeft kan passeren.
iii) Het DNA-bindende domein (DBD), dat zorgt voor de binding van het ligand-receptorcomplex aan een
specifiek stukje DNA, het Androgeen Respons Element (ARE).
iv) Het transactiveringsdomein (NTD), dat helpt bij de binding van een aantal begeleidende eiwitten. Die
eiwitten helpen bij het binden van het receptorcomplex met het ARE van het DNA. Ze helpen ook met kopieren
van het stuk DNA met de code voor het te synthetiseren eiwit.
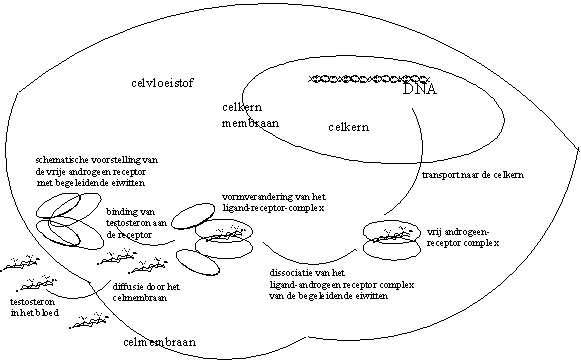
De lege androgeenreceptor is niet in vrije vorm aanwezig in de celvloeistof. Hij is geassocieerd met een aantal andere eiwitten. Nadat testosteron of dihydrotestosteron zich genesteld heeft in de holte in het Ligand Bindend Domein van de androgeen receptor, vindt een aanzienlijke vormverandering plaats van het complex als geheel. We kunnen deze vormverandering vergelijken met de vorming van een vuist om een voorwerp (zie Figuur 1).
Deze vormverandering heeft tot gevolg dat het ligand-receptorcomplex zich kan vrijmaken van zijn begeleidende eiwitten (zie Figuur 1). Hierdoor komen nieuwe plaatsen vrij aan de buitenkant van het ligand-receptorcomplex. Daardoor kunnen nieuwe interacties optreden.
Figuur 1
Klik voor groter figuur
Een van die nieuwe vrijgekomen plaatsen is het kern-translocatie-signaal (NLS). Dat zorgt ervoor dat het ligand-receptorcomplex het membraan dat de celkern omgeeft kan passeren. Vrij testosteron kan overigens ook het membraan dat de celkern omgeeft passeren, en daarna binden met lege androgeenreceptoren in de celkern.
Een andere vrijgekomen plaats op het getransformeerde complex van receptor en androgeen zorgt ervoor dat het complex gaat dimeriseren. Dat wil zeggen dat twee moleculen van het complex met elkaar gaan associeren. Al deze interacties en associaties berusten op het soort interactie op moleculair niveau dat in hoofdstuk 6 beschreven is. We hebben het dan dus over dipool-dipool-interacties, waterstofbruggen en Van der Waalskrachten.
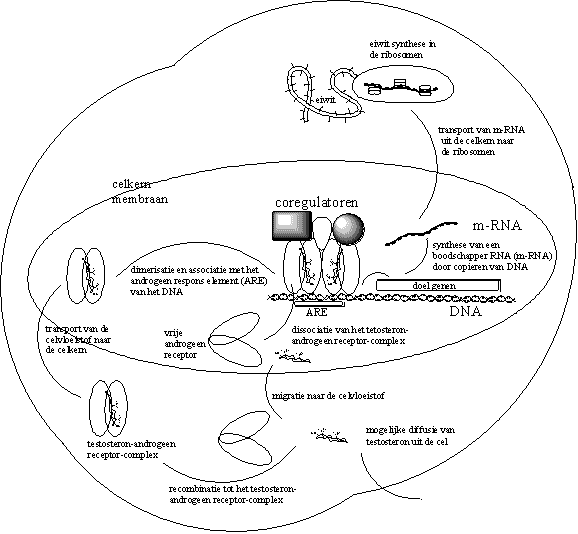
Het ligand-androgeenreceptor-dimeer gaat daarna complexeren met een code-element op het DNA. Deze code heet het Androgeen Respons Element (ARE). De associatie van het ligand-androgeenreceptor-dimeer met dat specifieke responselement geeft de aanzet tot kopieren van een in de buurt liggend stuk DNA. Dat bevat op zijn beurt de code voor het te synthetiseren eiwit (zie Figuur 2).
Figuur 2
Klik voor groter figuur
Het kopieren van het juiste stuk DNA gaat niet vanzelf. Er zijn een aantal enzymen voor nodig. In het volgende hoofdstuk vertellen we meer over de werking van enzymen. Bovendien is er ook sturing voor dat proces nodig. Dat is werk voor een aantal coregulatoren (zie Figuur 2).
Een van die enzymen is een RNA-polymerase. Met behulp van dat enzym kopieert de cel het juiste deel van het DNA. Zo synthetiseert de cel een boodschapper-RNA (een messenger-RNA of m-RNA). Boodschapper-RNA bevat een kopie van de genetische code van het DNA die codeert voor het te synthetiseren eiwit.
De cel transporteert het boodschapper-RNA vervolgens uit de celkern naar de ribosomen in de celvloeistof, waar de eiwitsynthese plaatsvindt. De ribosomen lezen de code van het boodschapper-RNA af, en vertalen die in de synthese van een nieuw eiwit. Dit nieuwe eiwit verzorgt dan uiteindelijk de functie waar het lichaam om vraagt. In het geval van bodybuilders is dat de opbouw van meer spierweefsel.
In de celvloeistof en in de celkern vindt ook voortdurend afbraak van eiwitten plaats. De wetenschappelijke term voor dit proces is proteolyse, ofwel splitsing van proteinen. De afbraak van eiwitten heeft tot gevolg dat het complex van eiwitten en receptor op het DNA op een gegeven moment desintegreert. De ligand komt dan weer vrij. Dit is het lot van de meeste steroid-receptorcomplexen.
Bij het testosteron-androgeenreceptorcomplex lijkt het iets anders te gaan. Het complex van androgeenreceptor en zijn ligand valt spontaan weer uit elkaar. De lege receptor en het testosteron migreren (verhuizen) uit de kern naar de celvloeistof en kunnen daar eventueel opnieuw een complex vormen.
Het vrije testosteron kan ook uit de cel diffunderen naar het bloed. De bloedbaan transporteert het hormoon naar de lever, waar enzymen het om kunnen zetten in een derivaat dat niet meer actief is, en dat uit het lichaam verdwijnt. Het is ook mogelijk dat het steroid de celvloeistof weer binnenkomt en met de receptor bindt. Er vindt dan een recirculatie plaats.
Het is duidelijk dat de boven beschreven processen berusten op een groot aantal delicate interacties. Alles luistert nauw, en het is niet gemakkelijk stoffen te vinden die al deze processen precies op dezelfde manier of beter stimuleren dan de natuurlijke steroidhormonen. De associatie- en dissociatieprocessen moeten op het juiste moment en in de juiste mate plaatsvinden. Een synthetisch anabool zal een net iets andere pasvorm in de receptorholte hebben, en dat zal op een net iets andere manier doorwerken in de vorm en associatiemogelijkheden van de verschillende domeinen.
De bijwerkingen van synthetische anabole steroďden zijn daardoor onvoorspelbaar. Het is niet mogelijk om op basis van een structuurformule die bijwerkingen te voorspellen.
Die onvoorspelbaarheid maakt uitvoerige biologische testen van nieuwe anabolen noodzakelijk. Het is bedenkelijk dat deze testen bij veel nieuwe steroiden die op de markt komen niet zijn uitgevoerd. Gebruikers van deze preparaten lopen daardoor extra gezondheidsrisico’s.
[1] T. Saartok, E. Dahlberg en J.A. Gustafson, Endocrinology (1984) 114, 2100-2106.
[2] E.T. Keller, W.B. Ershler en C. Chang, Frontiers in Bioscience (1996) 1, d59-71.
[3] J. Gobinet, N. Pujol en Ch. Sultan, Molecular and Cellular Endocrinology (2002) 198, 15-24.
[4] H.-J. Lee en C.Chang, Cellular and Molecular Life Sciences (2003) 60, 1613-1622.
[5] D.K. Lee en C. Chang, The Journal of Clinical Endocrinology and Metabolism (2003) 88, 4043-4054.
[6] W. Gao, C.E Bohl en J.T. Dalton, Chemical Reviews (2005) 105, 3352-3370.
|
|